
Genome-wide association analysis for yield and potassium efficiency related traits of wheat at maturity stage
-
摘要:目的
探明不同钾供应条件下控制产量及钾效率相关性状的稳定的显著关联分子标记位点,为小麦产量及钾效率相关性状的遗传控制机理研究及相关基因的克隆提供参考。
方法利用134个小麦品种 (系) 组成的群体为试验材料,设置正常供钾 (T1) 和不施钾 (T2) 两个处理,进行了2年田间试验 (E1、E2)。对小麦成熟期株高、穗长、穗粒数及钾吸收、利用效率等23个性状进行了表型鉴定,分别定义了同年同一处理和同一处理两年平均共6个环境平均值。采用GLM+Q一般线性模型和MLM+K+Q混合线性模型相结合的方法,利用群体差异SNP分子标记 (90K SNP芯片) 对小麦产量和钾效率相关性状进行全基因组关联分析。
结果与正常钾处理相比,不施钾处理条件下籽粒钾利用效率显著升高,单株钾累积量、单株钾含量及总小穗数等性状显著降低。供试小麦各性状的群体变异系数为6.98%~350.38%,有14个性状的遗传力在50%以上,以株高的遗传力最高 (92.03%)。利用保留的7485个多态性好的群体差异 (SNP) 进行了全基因组关联分析,共检测到1420个分子标记位点与供试23个性状在P ≤ 0.001水平存在显著关联,分布在21条染色体上。有1097个 (77.25%) 分子标记位点仅在一个关联分析环境中被检测到;能在至少两个关联分析环境中被检测到的相对稳定分子标记位点有323个,其中113个位点与钾效率相关性状有关,Tdurum_contig26281_139、Kukri_c307_2053等分子标记位点可以提高钾吸收效率,Ex_c19038_571、BS00039148_51等分子标记位点能够提高钾利用效率。在至少4个关联分析环境中被检测到的位点有22个,分别与株高、千粒重、穗粒数等5个性状相关。与株高和千粒重显著关联的分子标记位点RFL_Contig4069_2628和BS00003632_51可同时在全部6个关联分析环境中检测到,平均贡献率为9.59%和13.66%,环境稳定性非常好,与株高的降低和千粒重的提高显著关联。
结论不同钾供应水平下与产量及钾效率相关性状显著关联的分子标记位点存在显著差异,77.25%的分子标记位点仅在特定环境下被检测到。但也有22个显著关联分子标记位点 (涉及9个产量及钾效率相关性状) 在至少4个关联分析环境 (共6个环境) 下被检测到,形成高频表达分子标记位点。其中与株高和千粒重分别显著关联的两个分子标记位点在所有6个关联分析环境中均稳定被检测到,能显著降低株高和提高千粒重。这些分子标记位点的相关基因对相关性状的调控效应受钾处理环境影响小,具有较高的理论和应用价值,值得深入研究。
Abstract:ObjectivesCrops are found response genetically to potassium (K) nutrition, so the identification and localization of the molecular markers, that are stable and significantly correlated to traits of yield and K efficiency under different K nutrition levels, will provide reference for further genetic modification of related traits and genes cloning in breeding.
MethodsA group of 134 wheat varieties (or lines) were used in a two-years field experiment, in which applying K (T1) and not applying K (T2) were setup. The yield and K efficiency related traits of wheat were measured, and the average data of traits were calculated under 6 defined environments, as the respective average values of each treatment in one year and two years. Genome-wide association analysis was carried out for the tested traits using SNP molecular markers.
ResultsCompared with K application treatment, the K use efficiencies were significantly higher, while the K contents and accumulation of individual plant and the total spikelet number were significantly lower under no K application treatment. The coefficients of variation of the traits were 6.98%–350.38% among the group of cultivars (lines), and 14 of traits had heritability of > 50%, with the highest trait of plant height (92.03%). According to genome-wide association analysis on the 7485 SNPs with satisfactory diversity, a total of 1420 molecular marker sites were identified associating with 23 traits significantly (P ≤ 0.001), and located in 21 chromosomes. There were 1097 (77.25%) molecular marker sites detected in only one environment, and 323 molecular marker sites that could be stably identified in at least 2 environments, and 113 of them were related to K efficiency, e.g. the molecular marker sites Tdurum_contig26281_139 and Kukri_c307_2053 could upregulate the absorption of K, Ex_c19038_571 and BS00039148_51 could increase K use efficiency. There were 22 molecular marker sites that could be identified in at least 4 environments, which were related to 5 traits such as plant height, 1000-grain weight (1000-GW) and grain number per spike. Among the 22 molecular marker sites, RFL_Contig 4069_2628 and BS00003632_51 were related to plant height and 1000-GW, and were extremely stable and could be detected in all the 6 environments. The expression of genes in the two molecular marker sites significantly affected the variation of plant height and 1000-GW, with average contribution rate of 9.59% and 13.66%.
ConclusionsUnder different K nutrition levels, the molecular markers that are closely correlated to the yield and K efficiency traits, are detected in different sites. About 77.25% of the molecular marker sites are detected only in specific environments. However, 22 significantly correlated molecular marker sites (involving 9 traits related to yield and K efficiency) can be detected in at least 4 environments (6 environments in total), forming high-frequency expression molecular marker sites. Among them, two molecular marker sites are stably correlated with plant height and 1000-GW in all environments. The regulation effects of related genes on related traits of these molecular marker sites are less affected by K treatments, which has high theoretical and applied value and is worthy of in-depth study.
-
Keywords:
- wheat /
- association analysis /
- yield /
- potassium efficiency
-
小麦是世界上最重要的粮食作物之一,在中国的种植面积超过2.45亿hm2,产量超过1.34亿t (http://faostat.fao.org)。钾是植物必需的大量营养元素之一,对提高作物产量及品质具有重要意义[1-2]。然而,全世界130亿hm2的土壤中,受矿质养分严重胁迫的占22.5%,其中大约40%的养分胁迫土壤缺钾[3]。世界上农业用地大面积缺钾,在过去的10年里,钾肥平均每年施用616万t (http://data.stats.gov.cn/easyquery.htm?cn=C01),同时,农产品的产出和土壤淋洗也加重了土壤钾的缺乏[4]。与此同时,我国钾矿资源缺乏,钾肥的进口依赖度超过50%,提高钾效率是小麦生产可持续发展的必然要求。发掘小麦中的钾高效基因,对小麦进行钾高效定向培育改良是提高小麦钾效率的重要途径。
对现有小麦品种进行钾效率评价,挖掘钾高效小麦资源是品种选育的前提和基础。钾效率通常被定义为在低钾胁迫下产量潜力所占的比例[3-5]。钾高效的基因型通常在缺钾土壤中表现出较高的钾吸收能力 (钾吸收效率)[6-7]或单位钾吸收量的干物质产量较高[8-9] 。利用产量、生物量相关指标和钾吸收累积及利用相关指标可以在一定程度上全面反映小麦的钾效率情况。通过关联分析可以找到与这些钾效率相关指标显著相关的分子标记,从而进一步挖掘相关基因。全基因组关联分析 (genome wide association study, GWAS) 是一种以不同基因座等位基因间的连锁不平衡 (LD) 为基础,在群体内进行分子标记与性状的相关性分析,来鉴定目标性状与染色体区段关系的分析方法[10-11]。关联分析基于连锁的不平衡,以自然群体为研究对象,检测到的标记基本位于前人定位的QTL位点所在区段,并且定位结果精度较高,因此关联分析是一种十分有效的标记定位方法。目前,通过关联分析研究,已经发现了许多与小麦产量、株高等农艺性状相关的标记位点并发掘了一些位点的优异等位变异[12-14],与小麦钾养分效率相关性状的QTL分析已有部分报道,也获得了一些相关分子标记位点。例如,Kong等[15]设置了不同钾处理下的水培、池栽和大田试验,在“Chuan 35050 × Shannong 483”群体中共定位到80个与小麦成熟期性状有关和87个与小麦苗期性状有关的QTL位点;宫晓平[16]利用SL (鲁麦21 × 山农0431) 群体在不同钾处理下定位到79个小麦产量性状的QTL。
由于SNP在基因组中数量多、分布广,具有较高的遗传稳定性,并且易实现自动化分析,在小麦遗传学上应用越来越广泛。本研究以134份小麦品种 (系) 组成的自然群体为研究材料,对施钾和不施钾两个处理下小麦成熟期产量性状和钾效率相关性状共23个性状进行全基因组关联分析,以期获得与这些性状显著关联的分子标记位点,为钾高效小麦育种和相关基因克隆提供参考和数据支持。
1. 材料与方法
1.1 试验材料
材料为源自山东、河北、河南、陕西、山西、江苏、四川、澳大利亚和加拿大的共134个小麦品种 (系) 组成的群体 (表1)。
表 1 供试小麦品种Table 1. Wheat varieties used in this study序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)1 昌乐5号 28 山农45 55 淄12 82 石麦15号 109 蓝58 2 德选1号 29 山农483 56 954(7)-8 83 石麦16号 110 陕627 3 济麦20 30 山农664 57 BSS 84 石麦18号 111 陕农138 4 济麦21 31 山农8355 58 LS3283 85 石新616 112 陕农534 5 济麦22 32 山农紫麦1 59 山农25 86 石新618 113 西农85 6 济南17 33 山融3号 60 LS4697 87 石新828 114 西农213 7 济宁16 34 泰农18 61 LS4942 88 石优17号 115 西农889 8 济宁17 35 泰山21 62 LS6045 89 唐麦8号 116 西农9871 9 科信9号 36 泰山22 63 山农29 90 小偃81 117 小偃22 10 莱州137 37 泰山23 64 M8008 91 洛旱6号 118 小偃92 11 莱州95021 38 泰山24 65 贵农35选21 92 洛旱8号 119 小偃216 12 良星99 39 郯麦98 66 薛/淮 93 洛旱21号 120 运丰139 13 临麦4号 40 潍麦7号 67 藁优9618 94 洛旱23号 121 黑小麦76 14 临麦6号 41 潍麦8号 68 邯00-7086 95 洛旱24号 122 晋麦33号 15 鲁麦14 42 汶农5号 69 河农827 96 漯珍1号 123 临丰3号 16 鲁麦21 43 汶农6号 70 河农4198 97 新麦16 124 临旱822 17 鲁麦22 44 鑫289 71 河农6049 98 新麦18 125 临抗16号 18 鲁麦23 45 烟5072 72 衡0628 99 新麦26 126 临优145 19 齐丰1号 46 烟农999 73 衡6599 100 豫麦49 127 品资旱99-2 20 山农0431 47 烟99603 74 冀5265 101 豫麦54 128 平阳298 21 山农1186 48 烟农0428 75 冀丰703 102 郑麦9023 129 淮麦16 22 山农12 49 烟农15 76 金麦1号 103 中优9507 130 淮麦18 23 山农15 50 烟农19 77 科农199 104 周99233 131 川35050 24 山农17 51 烟农21 78 廊研43号 105 周黑麦1号 132 中国春 25 山农18 52 烟农22 79 石54 106 周麦24 133 澳-红麦 26 山农21 53 烟农23 80 石4185 107 AN2 134 加-超强筋麦 27 山农23 54 烟农836 81 石家庄8号 108 AN3 注(Note):表中品种 (系) 来源 Source of cultivar (line) listed in the table: 1~66, 中国山东 Shandong, China; 67~74, 中国山西 Shanxi, China; 75~90, 中国河南 Henan, China; 91~114, 中国河北 Hebei, China; 115~128, 中国陕西 Shaanxi, China; 129~130, 中国江苏 Jiangsu, China; 131~132, 中国四川 Sichuan, China; 133, 澳大利亚 Australia; 134, 加拿大Canada. 1.2 试验设计
1.2.1 试验处理
试验设在山东省泰安市山东农业大学农学实验站,试验时间为2012年10月7日至2013年6月9日 (E1) 和2013年10月6日至2014年6月8日 (E2)。供试土壤为褐土,pH 7.8,土壤速效氮、磷、钾含量分别约为84、25和82 mg/kg。试验设施钾 (T1) 和不施钾 (T2) 两个处理,所有处理小麦均施尿素N 195 kg/hm2、P2O5 92 kg/hm2 (过磷酸钙),施钾小区施K2O 114 kg/hm2 (硫酸钾),每个处理重复两次,随机排列。同一个钾处理小区内,小麦品种随机排列,排列位置两年相同。
1.2.2 试验方案
按株距10 cm、行距23 cm点播,行长40 cm,每个材料种植3行,每行点播15粒种子,每行留苗5株。磷、钾肥作为基肥一次施用,氮肥按照基肥∶拔节肥=5∶5的比例施用,正常田间管理。
1.2.3 性状调查
收获前10天分别对每个材料中间一行5个单株的总穗数 (SN)、主茎株高 (PH)、穗长 (SL)、基部不育小穗数 (BSSS)、顶部不育小穗数 (TSSS)、总小穗数 (TSSN) 和穗粒数 (GNPS) 进行调查统计,计算可育小穗数 (FSS)、不育小穗数 (SSS),取平均值代表该材料的相关性状。收获时,将各材料选取的中间一行5个单株地上部收获至网袋中,及时晾晒。晒干后称量地上部总重获得地上部总生物量后脱粒,脱粒时将颖壳收集起来与秸秆合并粉碎为秸秆样品,留样测定。称取籽粒总重,计算单株籽粒产量 (GWP)、单株秸秆产量 (StWP) 和单株地上部总生物量 (SWP),其中StWP=SWP–GWP。
1.2.4 测定方法
植株籽粒钾含量 (GKC)、秸秆钾含量 (StKC) 采用H2SO4-H2O2消解—火焰光度法测定,地上部钾含量 (SKC) = (GKC × GWP+StKC × StWP)/SWP;钾累积量 (籽粒钾累积量GKA、秸秆钾累积量StKA) 为相应部位的钾含量与其生物量的乘积,地上部钾累积量 (SKA)=GKA+StKA;钾利用效率为单位钾含量支撑的生物量[17],钾利用效率=生物量/该部位钾含量;收获指数 (HI)=GWP/SWP;千粒重 (1000-GW) 通过每个材料随机选择3个200粒称重换算。
1.3 表型数据分析
利用SPSS 17.0软件对小麦成熟期产量及钾效率相关性状进行表型数据分析。性状广义遗传力计算公式[18]为:hB2 = σg2/(σg2+σe2),式中,σg2和σe2分别为遗传方差和环境方差。
1.4 SNP标记检验及群体结构分析
使用美国Illumina公司和美国堪萨斯州立大学共同开发的小麦90K SNP芯片进行群体的基因组分析,共检测到91829个SNP标记,覆盖了小麦全基因组[19]。通过对获得的差异标记信息进行单拷贝SNP筛选,去除最小等位基因频率 (MAF) 小于5%、缺失率大于15%的SNP位点,保留7485个多态性好的SNP[20]。利用Powermarker 3.25软件分析等位变异数并计算多态性信息含量 (polymorphism information content, PIC)。多态性信息含量的计算公式为:
PIC=1−i∑j=1P2ij ,式中,Pij为位点i的第 j个等位变异出现的频率。在每条染色体每隔15 cM选取一个SNP,保证两个SNP间互相独立,最终用375个标记和软件Structure 2.3.4进行群体结构分析[20-21]。将群体数目K值设为1~12,每个K值重复5次,并假设位点间是相互独立的。开始时的不作数迭代 (length of burnin period) 设为10000,不作数迭代后的MCMC (Markov Chain Monte Carlo) 设为100000,然后参照Evanno等[22]的方法确定亚群数目K。同时获得Q矩阵作为关联分析中的协方差矩阵。利用软件PowerMarker V3.25 分析SNP的PIC值、基因多样性和遗传距离[20]。
1.5 数据分析
采用TASSEL 5.0软件,利用GLM+Q (general linear model) 一般线性模型和MLM+K+Q (mixed linear model) 混合线性模型结合群体分子标记数据和表型数据对两年两个钾处理 (T1E1、T2E1、T1E2、T2E2) 以及相同钾处理两年平均值 (T1AV、T2AV) 共6个关联分析环境下进行关联分析,确定显著关联分子标记 (P ≤ 0.001)。
2. 结果与分析
2.1 不同钾供应条件下供试群体产量、钾效率相关性状及其遗传力分析
表2表明,与施钾处理相比,不施钾处理小麦产量相关性状穗长、总小穗数、地上部生物量、单株钾累积量和钾含量均显著下降,收获指数和籽粒钾利用效率显著升高。不同基因型之间差异显著,供试群体变异系数6.98%~350.38%。
表 2 产量性状及钾效率相关性状的统计及遗传力分析Table 2. Statistical and heritability analysis for yield traits and K efficiency related traits性状
Trait处理
Treatment测定值 (AV ± SD)
Measured value遗传力(%)[18]
Heritability性状
Traits处理
Treatment测定值 (AV ± SD)
Measured value遗传力 (%)[18]
Heritability穗数
Spike numberT1E1 12.80 ± 3.13 a 52.85 秸秆产量
Straw weight
(g/plant)T1E1 28.69 ± 6.14 a 53.96 T2E1 11.57 ± 3.15 a T2E1 24.29 ± 7.09 b T1E2 10.43 ± 2.77 a T1E2 29.12 ± 9.54 a T2E2 8.80 ± 2.42 b T2E2 25.04 ± 8.41 b 株高
Plant height
(cm)T1E1 64.32 ± 9.50 a 92.03 地上部生物量
Shoot weight
(g/plant)T1E1 52.58 ± 10.29 a 55.25 T2E1 58.41 ± 8.89 b T2E1 44.47 ± 11.73 b T1E2 73.01 ± 11.79 a T1E2 52.63 ± 13.57 a T2E2 70.80 ± 11.59 a T2E2 45.93 ± 11.69 b 穗长
Spike length
(cm)T1E1 9.88 ± 1.26 a 60.83 籽粒钾含量
Grain K content
(g/kg)T1E1 5.23 ± 0.67 a 71.60 T2E1 9.46 ± 1.21 b T2E1 3.74 ± 0.74 b T1E2 9.85 ± 2.88 a T1E2 4.75 ± 0.57 a T2E2 9.28 ± 1.16 b T2E2 4.39 ± 0.67 b 总小穗数
TSSNT1E1 19.40 ± 1.35 a 80.37 秸秆钾含量
Straw K content
(g/kg)T1E1 29.99 ± 4.85 a 4.83 T2E1 18.77 ± 1.50 b T2E1 16.49 ± 4.82 b T1E2 19.18 ± 1.42 a T1E2 20.60 ± 3.01 a T2E2 19.20 ± 1.37 b T2E2 17.58 ± 3.21 b 基部不育
小穗数
BSSST1E1 0.44 ± 0.43 a 46.75 地上部钾含量
Shoot K content (g/kg)T1E1 35.13 ± 4.92 a 4.25 T2E1 0.37 ± 0.37 a T2E1 20.52 ± 4.78 b T1E2 0.62 ± 0.47 a T1E2 25.35 ± 3.13 a T2E2 0.51 ± 0.52 b T2E2 21.96 ± 3.29 b 顶部不育
小穗数
TSSST1E1 0.35 ± 0.51 a 41.60 籽粒钾累积量
Grain K accumulation
(g/plant)T1E1 0.13 ± 0.03 a 45.45 T2E1 0.31 ± 0.51 a T2E1 0.08 ± 0.03 b T1E2 0.04 ± 0.15 a T1E2 0.11 ± 0.03 a T2E2 0.09 ± 0.24 a T2E2 0.09 ± 0.03 b 不育小穗数
SSST1E1 0.79 ± 0.67 a 51.86 秸秆钾累积量
Straw K accumulation
(g/plant)T1E1 0.86 ± 0.21 a 16.34 T2E1 0.68 ± 0.70 b T2E1 0.41 ± 0.19 b T1E2 0.65 ± 0.49 a T1E2 0.60 ± 0.21 a T2E2 0.59 ± 0.56 a T2E2 0.43 ± 0.15 b 不同性状的遗传力存在显著差异,有14个性状遗传力较高,在50%以上,其中株高的遗传力最大 (92.03%),地上部钾含量的遗传力最小 (4.25%)。株高在不同钾处理下仍然主要受遗传因素控制,遗传力较高,而地上部钾含量受钾处理影响大,遗传力较小。
2.2 产量及钾效率相关性状的分子标记及稳定表达的分子标记位点
将23个产量及钾效率相关性状分别与两年两个钾处理 (T1E1、T2E1、T1E2、T2E2) 以及相同钾处理两年平均值 (T1-AV、T2-AV) 共6个关联分析环境的供试性状进行分子标记关联分析,共检测到1420个关联极显著 (P ≤ 0.001) 的分子标记位点,分布在21条染色体上。关联分析结果中,R2为特定表型性状 (测定值) 在不同钾处理环境下产生的变异 (表型变异) 由该分子标记/基因贡献的比例 (贡献率)。本研究检测到的显著关联分子标记位点对相应性状表型变异的贡献率 (R2) 范围是7.53%~18.01% (表3)。其中,与千粒重 (1000-GW) 关联显著的位点最多,共269个,表型变异贡献率为7.80%~18.01% (R2);与籽粒钾含量 (GKC) 关联的位点最少,只定位到23个,表型变异贡献率7.74%~13.51% (R2)。在定位到的全部1420个显著关联的分子标记位点中,有1097个分子标记位点仅在一个关联分析环境中被检测到,占总数的77.25%。有323个分子标记位点至少在两个关联分析环境中被检测到,其中113个位点与钾效率相关性状有关,Tdurum_contig26281_139、Kukri_c307_2053等分子标记位点可以提高钾吸收效率,Ex_C19038_571、BS00039148_51等分子标记位点能够提高钾利用效率。其中有22个分子标记位点至少在四个关联分析环境中被检测到,分别与株高、千粒重、穗粒数等5个性状相关,形成高频表达分子标记位点 (表4)。值得注意的是,分别与株高和千粒重显著关联的分子标记位点RFL_Contig4069_2628和BS00003632_51在全部6个关联分析环境中均被检测到,平均贡献率分别为9.59%和13.66%,环境稳定性非常好 (表4)。
表 3 与产量及钾效率相关性状显著相关的分子标记数及其对性状表型变异的贡献率Table 3. Markers significantly associated with yield and K efficiency traits and their contribution to phenotypic variations性状
Trait定位分子标记数
Located markers稳定标记数
Stable markers贡献率 (%)
R2最大R2标记
Max-R2 marker最大R2位置
Max-R2 site标记在染色体上的分布
Markers distributionPH 32 18 7.85~11.87 RFL_Contig5170_330 6A 1A,2A,2B,3A,3B,4B,5A,5B,6A,6B,7A SL 53 5 7.99~13.72 BobWhite_c1456_615 1B 1A,1B,3A,4B,4D,5A,5D,6A,7A,7B SN 66 6 7.97~13.37 BS00029013_51 1B 1B,2B,2D,3B,4A,4B,5A,5D,6B,7A,7D BSSS 37 10 7.53~12.87 Tdurum_contig70123_354 5B 1A,2B,2D,3B,4A,4B,5A,5B,6B,7A TSSS 35 3 7.69~11.89 Kukri_c39767_204 5B 1A,1B,4B,5B,6A,6B,7A,7B FSS 24 4 7.56~14.68 Tdurum_contig43078_90 5B 1A,1B,2B,3B,4A,4B,5A,5B,6B,7A,7B SSS 25 1 8.13~13.05 Excalibur_c25239_283 3A 2D,3A,5A,6B TSSN 34 3 8.14~13.19 Excalibur_c25239_283 3A 1B,2A,2D,3A,3B,4A,5A,6B,7A GNPS 24 4 8.00~12.24 RAC875_c58966_471 5A 2A,2B,4B,5A,5B,6A,7B 1000-GW 269 137 7.80~18.01 BS00003632_51 6A 1A,1B,2A,2B,2D,3A,3B,3D,4B,5A,5B,5D,6A,6B,6D,7A HI 68 4 7.73~15.53 wsnp_Ex_c24135_33382521 2B 1A,1B,2B,3A,3B,3D,4A,4B,5A,5B,6A,7A,7B GWP 39 2 7.82~12.05 Excalibur_c46601_265 7A 1B,1D,2B,2D,3B,5B,6A,6B,7A,7D StWP 33 7 7.56~12.56 BobWhite_c3506_1151 6B 1B,2D,3A,3B,5A,5B,6A,6B,6D,7B SWP 25 5 7.90~12.54 wsnp_Ex_c8208_13870372 3B 3A,3B,6A,6B,7A,7D GKA 91 12 7.89~13.57 Kukri_c13136_1363 1A 1A,1B,2A,2B,2D,3A,3B,4A,4B,5B,6A,6B,7A,7B,7D StKA 127 25 7.89~13.02 Tdurum_contig7992_605 4A 1A,1B,1D,2A,2B,2D,3A,4A,5A,5B,6A,6B,6D,7A,7D SKA 139 26 7.90~13.46 Tdurum_contig7992_605 4A 1A,1B,1D,2A,2B,2D,3A,3B,3D,4A,4B,5A,5D,6A,6B,6D,7A,7B,7D GKC 23 2 7.74~13.51 Excalibur_c46601_265 7A 1D,2D,4A,5A,5B,6A,7A,7D StKC 80 19 7.68~13.47 CAP7_c3178_52 3A 1B,2A,2B,2D,3A,3B,3D,4A,5A,5B,6A,6B,7A,7B SKC 60 14 8.00~13.50 CAP7_c3178_52 3A 1B,2A,2B,2D,3A,3B,3D,4A,5B,5D,6A,6B,7A,7B GKUE 40 4 7.90~11.71 wsnp_Ex_c8208_13870372 3B 1B,2A,2B,3B,4A,5B,6A,7A,7B,7D StKUE 64 9 7.58~14.25 CAP7_c4283_67 6A 1A,1B,1D,2A,2B,3A,3B,4A,4B,5A,5B,5D,6A,6B,6D,7A,7B SKUE 32 3 7.82~12.81 wsnp_Ex_c8208_13870372 3B 1B,2B,3B,4A,4B,5A,5B,6A,7A 注(Note):PH—株高 Plant height; SL—穗长 Spike length; SN—穗数 Spike number; BSSS—基部不育小穗数 Base sterile small spikes; TSSS—顶部不育小穗数 Top sterile small spikes; FSS—可育小穗数 Fertile small spikes; SSS—不育小穗数 Sterile small spikes; TSSN—总小穗数 Total small spike number; GNPS—穗粒数 Grain number per spike; 1000-GW—千粒重 1000-grain weight; HI—收获指数 Harvest index; GWP—籽粒产量 Grain weight; StWP—秸秆产量 Straw weight; SWP—地上部生物量 Shoot weight; GKC—籽粒钾含量 Grain K content; StKC—秸秆钾含量 Straw K content; SKC—地上部钾含量 Shoot K content; GKA—籽粒钾累积量 Grain K accumulation; StKA—秸秆钾累积量 Straw K accumulation; SKA—地上部钾累积量 Shoot K accumulation; GKUE—籽粒钾利用效率 Grain K utilization efficiency; StKUE—秸秆钾利用效率 Straw K utilization efficiency; SKUE—地上部钾利用效率 Shoot K utilization efficiency. 表 4 稳定表达的产量和钾效率相关性状的标记及其关联标记位点Table 4. The environmental stable markers significantly associated with yield and K efficiency related traits性状
Trait标记
Markers染色体
Chr.位置 (cM)
LociP值 P-value 贡献率 R2 (%) T1AV T1E1 T1E2 T2AV T2E1 T2E2 T1AV T1E1 T1E2 T2AV T2E1 T2E2 GKA RAC875_c52504_215 6A 45.11 8.08E-04 7.04E-05 8.59E-04 6.56E-05 8.20 11.30 8.10 11.31 GNPS BS00060796_51 1A 25.34 6.06E-04 9.92E-04 7.03E-04 5.32E-04 8.42 7.88 8.26 8.59 PH Excalibur_c15332_1194 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 RAC875_rep_c115516_134 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 Excalibur_c15332_453 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 RFL_Contig5170_330 6A 16.58 3.94E-05 6.13E-05 1.11E-04 5.14E-04 8.66E-05 11.87 11.44 10.60 8.71 10.89 wsnp_Ku_c1468_2912489 6A 138.04 4.62E-04 8.47E-04 3.32E-04 6.74E-04 8.77 8.08 9.21 8.29 RFL_Contig4069_2628 6A 140.87 2.95E-04 4.82E-04 5.76E-04 9.04E-05 3.31E-04 1.08E-04 9.33 8.81 8.51 10.93 9.36 10.61 RAC875_c28630_214 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 RAC875_c7828_598 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 RAC875_c42866_1323 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 BS00046264_51 6B 109.70 9.21E-04 8.09E-04 3.91E-04 1.60E-04 7.62 7.89 9.13 10.33 StWP wsnp_Ra_c33025_41968284 1A 82.67 9.11E-04 5.38E-04 2.28E-04 5.04E-04 0.001 7.96 8.68 9.66 8.66 7.83 1000-GW Kukri_c29039_315 1A 83.70 4.88E-04 7.29E-04 1.04E-04 1.14E-04 9.75E-04 8.77 8.29 10.67 10.54 7.87 GENE-4064_599 3B 61.64 4.32E-04 8.94E-06 2.63E-05 8.04E-05 8.92 13.72 12.37 11.05 Excalibur_c58066_180 3B 60.25 9.44E-04 4.50E-05 9.72E-05 3.19E-04 7.92 11.71 10.74 9.30 BS00023193_51 3B 59.78 9.44E-04 4.50E-05 9.72E-05 3.19E-04 7.92 11.71 10.74 9.30 wsnp_Ku_c4078_7436510 3B 71.34 6.85E-04 2.43E-04 6.69E-04 8.69E-04 8.33 9.59 8.30 8.02 wsnp_Ku_c21818_31604716 3B 71.34 6.85E-04 2.43E-04 6.69E-04 8.69E-04 8.33 9.59 8.30 8.02 RAC875_c16943_404 5A 80.43 2.06E-04 2.95E-05 7.65E-04 8.72E-04 9.87 12.38 8.13 7.96 BS00003632_51 6A 22.01 3.29E-06 5.17E-04 3.28E-04 2.54E-07 6.76E-07 7.40E-06 15.05 8.73 9.30 18.01 16.84 14.04 BS00078460_51 7A 82.95 1.75E-05 1.71E-04 5.89E-04 7.90E-04 12.98 10.14 8.46 8.08 注(Note):GKA—籽粒钾累积量 Grain K accumulation; GNPS—穗粒数 Grain number per spike; pH—株高 Plant height; StWP—秸秆产量 Straw weight; 1000-GW—千粒重 1000-grain weight. 处理中, E1 和 E2 代表两个试验年份, T1、T2 代表施钾和不施钾处理, T1AV 和 T2AV 代表两年处理平均值 In the treatments, E1 and E2 represent the two experimental years, T1 and T2 represent K and no K application treatments, T1AV and T2AV represent the two years' averages of T1 and T2 treatments. 3. 讨论
3.1 施钾对小麦产量及钾效率相关性状的影响
钾是植物的大量营养元素之一,农作物对钾的需求仅次于氮,而土壤中常因钾供应不足影响作物产量。钾具有增强植物细胞抵御外界恶劣环境的能力,如高温、低温、干旱、倒伏等,低钾胁迫会限制植物生长,降低生物量[23]。本研究结果也发现,在两年不施钾条件下,单株钾累积量和单株钾含量显著低于施钾处理,但籽粒钾利用效率显著升高,表明在低钾胁迫条件下,小麦籽粒能够更加有效地利用钾元素[24]。本研究中不施钾处理还显著降低了小麦产量相关性状穗长、总小穗数。张会民等[2]的研究结果也表明,在缺钾处理下,小麦穗数、穗粒数和千粒重均显著低于施钾处理,施用钾肥对冬小麦的增产效果显著。
3.2 小麦产量和钾效率相关性状的显著关联位点
本研究采用GLM+Q一般线性回归和MLM+Q+K混合线性模型相结合的方法,并采用较高的阈值P
≤ 0.001,可有效消除由群体分层和亲缘关系引起的伪关联,增加关联结果的可信度。本研究利用全基因组7485个SNP标记与两年两个钾处理 (T1E1、T2E1、T1E2、T2E2) 以及相同钾处理两年平均值 (T1AV、T2AV) 共6个关联分析环境中小麦成熟期产量及钾效率相关性状进行了关联分析,共定位到1420个显著关联分子标记位点,分布在21条染色体上。所有检测到的显著关联分子标记表型变异贡献率 (R2) 范围是7.53%~18.01% (表3),表明所有供试性状均由多个微效基因控制,检测到的分子标记位点表型变异贡献率均不高。但是我们也发现有许多分子标记位点同时与多个性状关联,这与经典的一因多效现象相符。例如,Sukumaran等[25]在5A染色体上检测到的BS00079989_51位点与产量相关,在本研究中与不孕小穗数相关;Shi等[26]在3D染色体上发现的Kukri_c18420_705位点在本研究中与地上部钾累积量相关联;Zanke等[27]在3B染色体上发现的与千粒重关联的Tdurum_contig59953_220位点,在本研究中与顶部不育小穗数关联。本研究也发现了一些同时与多个钾效率相关性状显著关联的位点,如在6A染色体上检测到的分子标记位点Excalibur_rep_c111263_307、BobWhite_c23839_119同时与钾含量、累积量及利用效率性状显著关联。 不同环境下检测出的分子标记位点不完全一致,这可能是小麦适应不同环境条件的表现,这也是由多基因控制的数量性状的突出特点。本研究中1097个分子标记位点仅被检测到一次,占所有分子标记位点的77.25%,表明钾处理对产量及钾效率相关性状控制基因的表达影响显著,绝大多数分子标记位点/基因仅在特定环境下表达,一旦环境变化,就会有大量的不同分子标记位点/基因产生适应性变化。由于这些分子标记位点/基因数目多,对表型变异的贡献率较低,受环境影响大,因此很难锁定重要的适应性相关分子标记位点/基因。但我们也检测到一些环境稳定性好的分子标记位点,在至少两个关联分析环境中被检测到的相对稳定的显著关联分子标记位点有323个,占总数的22.75%。在相对稳定的显著关联分子标记位点中,有113个位点与钾效率相关性状有关,其中,Tdurum_contig26281_139、Kukri_c307_2053等分子标记位点与钾吸收效率的提高显著关联,Ex_c19038_571、BS00039148_51等分子标记位点能够显著提高钾利用效率。一些环境稳定性好的分子标记位点值得进一步关注,如至少在4个关联分析环境中被检测到的22个分子标记位点 (表4),还有两个分子标记位点RFL_Contig4069_2628和BS00003632_51分别与株高的降低和千粒重的提高相关联,在本试验检测的全部6个关联分析环境中均能定位到,平均表型变异贡献率分别为9.59%和13.66%。这些分子标记位点在多个钾处理环境中均能检测到,表明这几个分子标记位点基因的表达受钾处理环境影响较小,在不同钾处理环境条件下均可以稳定地与相关性状关联,形成高频表达分子标记位点。这些高频表达分子标记位点与众多QTL研究中关于高频QTL的研究结果[17-18]一致,这些分子标记位点相关基因可能更倾向于组成型表达,对相关性状调控效应的环境稳定性更强,因而具备更加实用的价值,值得深入挖掘与研究。
4. 结论
不同钾供应水平下与产量及钾效率相关性状显著关联的分子标记位点存在显著差异,77.25%的分子标记位点仅在特定环境下被检测到。但有22个显著关联分子标记位点(涉及9个产量及钾效率相关性状)在至少4个关联分析环境(共6个环境)下被检测到,形成高频表达分子标记位点。其中,与株高和千粒重分别显著关联的两个分子标记位点在所有6个关联分析环境中均稳定地被检测到,能在不同钾处理环境下稳定发挥作用,且显著降低株高和提高千粒重,这值得深入研究。
-
表 1 供试小麦品种
Table 1 Wheat varieties used in this study
序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)序号
No.品种 (系)
Cultivar (line)1 昌乐5号 28 山农45 55 淄12 82 石麦15号 109 蓝58 2 德选1号 29 山农483 56 954(7)-8 83 石麦16号 110 陕627 3 济麦20 30 山农664 57 BSS 84 石麦18号 111 陕农138 4 济麦21 31 山农8355 58 LS3283 85 石新616 112 陕农534 5 济麦22 32 山农紫麦1 59 山农25 86 石新618 113 西农85 6 济南17 33 山融3号 60 LS4697 87 石新828 114 西农213 7 济宁16 34 泰农18 61 LS4942 88 石优17号 115 西农889 8 济宁17 35 泰山21 62 LS6045 89 唐麦8号 116 西农9871 9 科信9号 36 泰山22 63 山农29 90 小偃81 117 小偃22 10 莱州137 37 泰山23 64 M8008 91 洛旱6号 118 小偃92 11 莱州95021 38 泰山24 65 贵农35选21 92 洛旱8号 119 小偃216 12 良星99 39 郯麦98 66 薛/淮 93 洛旱21号 120 运丰139 13 临麦4号 40 潍麦7号 67 藁优9618 94 洛旱23号 121 黑小麦76 14 临麦6号 41 潍麦8号 68 邯00-7086 95 洛旱24号 122 晋麦33号 15 鲁麦14 42 汶农5号 69 河农827 96 漯珍1号 123 临丰3号 16 鲁麦21 43 汶农6号 70 河农4198 97 新麦16 124 临旱822 17 鲁麦22 44 鑫289 71 河农6049 98 新麦18 125 临抗16号 18 鲁麦23 45 烟5072 72 衡0628 99 新麦26 126 临优145 19 齐丰1号 46 烟农999 73 衡6599 100 豫麦49 127 品资旱99-2 20 山农0431 47 烟99603 74 冀5265 101 豫麦54 128 平阳298 21 山农1186 48 烟农0428 75 冀丰703 102 郑麦9023 129 淮麦16 22 山农12 49 烟农15 76 金麦1号 103 中优9507 130 淮麦18 23 山农15 50 烟农19 77 科农199 104 周99233 131 川35050 24 山农17 51 烟农21 78 廊研43号 105 周黑麦1号 132 中国春 25 山农18 52 烟农22 79 石54 106 周麦24 133 澳-红麦 26 山农21 53 烟农23 80 石4185 107 AN2 134 加-超强筋麦 27 山农23 54 烟农836 81 石家庄8号 108 AN3 注(Note):表中品种 (系) 来源 Source of cultivar (line) listed in the table: 1~66, 中国山东 Shandong, China; 67~74, 中国山西 Shanxi, China; 75~90, 中国河南 Henan, China; 91~114, 中国河北 Hebei, China; 115~128, 中国陕西 Shaanxi, China; 129~130, 中国江苏 Jiangsu, China; 131~132, 中国四川 Sichuan, China; 133, 澳大利亚 Australia; 134, 加拿大Canada.  下载: 导出CSV
下载: 导出CSV
表 2 产量性状及钾效率相关性状的统计及遗传力分析
Table 2 Statistical and heritability analysis for yield traits and K efficiency related traits
性状
Trait处理
Treatment测定值 (AV ± SD)
Measured value遗传力(%)[18]
Heritability性状
Traits处理
Treatment测定值 (AV ± SD)
Measured value遗传力 (%)[18]
Heritability穗数
Spike numberT1E1 12.80 ± 3.13 a 52.85 秸秆产量
Straw weight
(g/plant)T1E1 28.69 ± 6.14 a 53.96 T2E1 11.57 ± 3.15 a T2E1 24.29 ± 7.09 b T1E2 10.43 ± 2.77 a T1E2 29.12 ± 9.54 a T2E2 8.80 ± 2.42 b T2E2 25.04 ± 8.41 b 株高
Plant height
(cm)T1E1 64.32 ± 9.50 a 92.03 地上部生物量
Shoot weight
(g/plant)T1E1 52.58 ± 10.29 a 55.25 T2E1 58.41 ± 8.89 b T2E1 44.47 ± 11.73 b T1E2 73.01 ± 11.79 a T1E2 52.63 ± 13.57 a T2E2 70.80 ± 11.59 a T2E2 45.93 ± 11.69 b 穗长
Spike length
(cm)T1E1 9.88 ± 1.26 a 60.83 籽粒钾含量
Grain K content
(g/kg)T1E1 5.23 ± 0.67 a 71.60 T2E1 9.46 ± 1.21 b T2E1 3.74 ± 0.74 b T1E2 9.85 ± 2.88 a T1E2 4.75 ± 0.57 a T2E2 9.28 ± 1.16 b T2E2 4.39 ± 0.67 b 总小穗数
TSSNT1E1 19.40 ± 1.35 a 80.37 秸秆钾含量
Straw K content
(g/kg)T1E1 29.99 ± 4.85 a 4.83 T2E1 18.77 ± 1.50 b T2E1 16.49 ± 4.82 b T1E2 19.18 ± 1.42 a T1E2 20.60 ± 3.01 a T2E2 19.20 ± 1.37 b T2E2 17.58 ± 3.21 b 基部不育
小穗数
BSSST1E1 0.44 ± 0.43 a 46.75 地上部钾含量
Shoot K content (g/kg)T1E1 35.13 ± 4.92 a 4.25 T2E1 0.37 ± 0.37 a T2E1 20.52 ± 4.78 b T1E2 0.62 ± 0.47 a T1E2 25.35 ± 3.13 a T2E2 0.51 ± 0.52 b T2E2 21.96 ± 3.29 b 顶部不育
小穗数
TSSST1E1 0.35 ± 0.51 a 41.60 籽粒钾累积量
Grain K accumulation
(g/plant)T1E1 0.13 ± 0.03 a 45.45 T2E1 0.31 ± 0.51 a T2E1 0.08 ± 0.03 b T1E2 0.04 ± 0.15 a T1E2 0.11 ± 0.03 a T2E2 0.09 ± 0.24 a T2E2 0.09 ± 0.03 b 不育小穗数
SSST1E1 0.79 ± 0.67 a 51.86 秸秆钾累积量
Straw K accumulation
(g/plant)T1E1 0.86 ± 0.21 a 16.34 T2E1 0.68 ± 0.70 b T2E1 0.41 ± 0.19 b T1E2 0.65 ± 0.49 a T1E2 0.60 ± 0.21 a T2E2 0.59 ± 0.56 a T2E2 0.43 ± 0.15 b
下载: 导出CSV
表 3 与产量及钾效率相关性状显著相关的分子标记数及其对性状表型变异的贡献率
Table 3 Markers significantly associated with yield and K efficiency traits and their contribution to phenotypic variations
性状
Trait定位分子标记数
Located markers稳定标记数
Stable markers贡献率 (%)
R2最大R2标记
Max-R2 marker最大R2位置
Max-R2 site标记在染色体上的分布
Markers distributionPH 32 18 7.85~11.87 RFL_Contig5170_330 6A 1A,2A,2B,3A,3B,4B,5A,5B,6A,6B,7A SL 53 5 7.99~13.72 BobWhite_c1456_615 1B 1A,1B,3A,4B,4D,5A,5D,6A,7A,7B SN 66 6 7.97~13.37 BS00029013_51 1B 1B,2B,2D,3B,4A,4B,5A,5D,6B,7A,7D BSSS 37 10 7.53~12.87 Tdurum_contig70123_354 5B 1A,2B,2D,3B,4A,4B,5A,5B,6B,7A TSSS 35 3 7.69~11.89 Kukri_c39767_204 5B 1A,1B,4B,5B,6A,6B,7A,7B FSS 24 4 7.56~14.68 Tdurum_contig43078_90 5B 1A,1B,2B,3B,4A,4B,5A,5B,6B,7A,7B SSS 25 1 8.13~13.05 Excalibur_c25239_283 3A 2D,3A,5A,6B TSSN 34 3 8.14~13.19 Excalibur_c25239_283 3A 1B,2A,2D,3A,3B,4A,5A,6B,7A GNPS 24 4 8.00~12.24 RAC875_c58966_471 5A 2A,2B,4B,5A,5B,6A,7B 1000-GW 269 137 7.80~18.01 BS00003632_51 6A 1A,1B,2A,2B,2D,3A,3B,3D,4B,5A,5B,5D,6A,6B,6D,7A HI 68 4 7.73~15.53 wsnp_Ex_c24135_33382521 2B 1A,1B,2B,3A,3B,3D,4A,4B,5A,5B,6A,7A,7B GWP 39 2 7.82~12.05 Excalibur_c46601_265 7A 1B,1D,2B,2D,3B,5B,6A,6B,7A,7D StWP 33 7 7.56~12.56 BobWhite_c3506_1151 6B 1B,2D,3A,3B,5A,5B,6A,6B,6D,7B SWP 25 5 7.90~12.54 wsnp_Ex_c8208_13870372 3B 3A,3B,6A,6B,7A,7D GKA 91 12 7.89~13.57 Kukri_c13136_1363 1A 1A,1B,2A,2B,2D,3A,3B,4A,4B,5B,6A,6B,7A,7B,7D StKA 127 25 7.89~13.02 Tdurum_contig7992_605 4A 1A,1B,1D,2A,2B,2D,3A,4A,5A,5B,6A,6B,6D,7A,7D SKA 139 26 7.90~13.46 Tdurum_contig7992_605 4A 1A,1B,1D,2A,2B,2D,3A,3B,3D,4A,4B,5A,5D,6A,6B,6D,7A,7B,7D GKC 23 2 7.74~13.51 Excalibur_c46601_265 7A 1D,2D,4A,5A,5B,6A,7A,7D StKC 80 19 7.68~13.47 CAP7_c3178_52 3A 1B,2A,2B,2D,3A,3B,3D,4A,5A,5B,6A,6B,7A,7B SKC 60 14 8.00~13.50 CAP7_c3178_52 3A 1B,2A,2B,2D,3A,3B,3D,4A,5B,5D,6A,6B,7A,7B GKUE 40 4 7.90~11.71 wsnp_Ex_c8208_13870372 3B 1B,2A,2B,3B,4A,5B,6A,7A,7B,7D StKUE 64 9 7.58~14.25 CAP7_c4283_67 6A 1A,1B,1D,2A,2B,3A,3B,4A,4B,5A,5B,5D,6A,6B,6D,7A,7B SKUE 32 3 7.82~12.81 wsnp_Ex_c8208_13870372 3B 1B,2B,3B,4A,4B,5A,5B,6A,7A 注(Note):PH—株高 Plant height; SL—穗长 Spike length; SN—穗数 Spike number; BSSS—基部不育小穗数 Base sterile small spikes; TSSS—顶部不育小穗数 Top sterile small spikes; FSS—可育小穗数 Fertile small spikes; SSS—不育小穗数 Sterile small spikes; TSSN—总小穗数 Total small spike number; GNPS—穗粒数 Grain number per spike; 1000-GW—千粒重 1000-grain weight; HI—收获指数 Harvest index; GWP—籽粒产量 Grain weight; StWP—秸秆产量 Straw weight; SWP—地上部生物量 Shoot weight; GKC—籽粒钾含量 Grain K content; StKC—秸秆钾含量 Straw K content; SKC—地上部钾含量 Shoot K content; GKA—籽粒钾累积量 Grain K accumulation; StKA—秸秆钾累积量 Straw K accumulation; SKA—地上部钾累积量 Shoot K accumulation; GKUE—籽粒钾利用效率 Grain K utilization efficiency; StKUE—秸秆钾利用效率 Straw K utilization efficiency; SKUE—地上部钾利用效率 Shoot K utilization efficiency.
下载: 导出CSV
表 4 稳定表达的产量和钾效率相关性状的标记及其关联标记位点
Table 4 The environmental stable markers significantly associated with yield and K efficiency related traits
性状
Trait标记
Markers染色体
Chr.位置 (cM)
LociP值 P-value 贡献率 R2 (%) T1AV T1E1 T1E2 T2AV T2E1 T2E2 T1AV T1E1 T1E2 T2AV T2E1 T2E2 GKA RAC875_c52504_215 6A 45.11 8.08E-04 7.04E-05 8.59E-04 6.56E-05 8.20 11.30 8.10 11.31 GNPS BS00060796_51 1A 25.34 6.06E-04 9.92E-04 7.03E-04 5.32E-04 8.42 7.88 8.26 8.59 PH Excalibur_c15332_1194 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 RAC875_rep_c115516_134 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 Excalibur_c15332_453 3B 73.35 5.38E-04 2.58E-04 3.42E-04 3.30E-04 8.57 9.61 9.23 9.20 RFL_Contig5170_330 6A 16.58 3.94E-05 6.13E-05 1.11E-04 5.14E-04 8.66E-05 11.87 11.44 10.60 8.71 10.89 wsnp_Ku_c1468_2912489 6A 138.04 4.62E-04 8.47E-04 3.32E-04 6.74E-04 8.77 8.08 9.21 8.29 RFL_Contig4069_2628 6A 140.87 2.95E-04 4.82E-04 5.76E-04 9.04E-05 3.31E-04 1.08E-04 9.33 8.81 8.51 10.93 9.36 10.61 RAC875_c28630_214 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 RAC875_c7828_598 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 RAC875_c42866_1323 7A 33.45 7.54E-04 7.38E-04 5.57E-04 3.89E-04 8.14 8.19 8.60 9.15 BS00046264_51 6B 109.70 9.21E-04 8.09E-04 3.91E-04 1.60E-04 7.62 7.89 9.13 10.33 StWP wsnp_Ra_c33025_41968284 1A 82.67 9.11E-04 5.38E-04 2.28E-04 5.04E-04 0.001 7.96 8.68 9.66 8.66 7.83 1000-GW Kukri_c29039_315 1A 83.70 4.88E-04 7.29E-04 1.04E-04 1.14E-04 9.75E-04 8.77 8.29 10.67 10.54 7.87 GENE-4064_599 3B 61.64 4.32E-04 8.94E-06 2.63E-05 8.04E-05 8.92 13.72 12.37 11.05 Excalibur_c58066_180 3B 60.25 9.44E-04 4.50E-05 9.72E-05 3.19E-04 7.92 11.71 10.74 9.30 BS00023193_51 3B 59.78 9.44E-04 4.50E-05 9.72E-05 3.19E-04 7.92 11.71 10.74 9.30 wsnp_Ku_c4078_7436510 3B 71.34 6.85E-04 2.43E-04 6.69E-04 8.69E-04 8.33 9.59 8.30 8.02 wsnp_Ku_c21818_31604716 3B 71.34 6.85E-04 2.43E-04 6.69E-04 8.69E-04 8.33 9.59 8.30 8.02 RAC875_c16943_404 5A 80.43 2.06E-04 2.95E-05 7.65E-04 8.72E-04 9.87 12.38 8.13 7.96 BS00003632_51 6A 22.01 3.29E-06 5.17E-04 3.28E-04 2.54E-07 6.76E-07 7.40E-06 15.05 8.73 9.30 18.01 16.84 14.04 BS00078460_51 7A 82.95 1.75E-05 1.71E-04 5.89E-04 7.90E-04 12.98 10.14 8.46 8.08 注(Note):GKA—籽粒钾累积量 Grain K accumulation; GNPS—穗粒数 Grain number per spike; pH—株高 Plant height; StWP—秸秆产量 Straw weight; 1000-GW—千粒重 1000-grain weight. 处理中, E1 和 E2 代表两个试验年份, T1、T2 代表施钾和不施钾处理, T1AV 和 T2AV 代表两年处理平均值 In the treatments, E1 and E2 represent the two experimental years, T1 and T2 represent K and no K application treatments, T1AV and T2AV represent the two years' averages of T1 and T2 treatments.
下载: 导出CSV
-
[1] Pettigrew W T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton[J]. Physiologia Plantarum, 2008, 133: 670–681. DOI: 10.1111/j.1399-3054.2008.01073.x
[2] 张会民, 刘红霞, 王林生, 等. 钾对旱地冬小麦后期生长及籽粒品质的影响[J]. 麦类作物学报, 2004, 24(3): 73–75. DOI: 10.3969/j.issn.1009-1041.2004.03.019 Zhang H M, Liu H X, Wang L S, et al. Effect of potassium on the growth at the late stages and grain quality of winter wheat in dry land[J]. Journal of Triticeae Crops, 2004, 24(3): 73–75. DOI: 10.3969/j.issn.1009-1041.2004.03.019
[3] 王利, 陈防, 万开元. 植物钾效率及其评价的研究进展与展望[J]. 土壤, 2010, 42(2): 164–170. Wang L, Chen F, Wan K Y. Progress and expectation of the research on plant K efficiency and its evaluation[J]. Soils, 2010, 42(2): 164–170.
[4] Rengel Z, Damon P M. Crops and genotypes differ in efficiency of potassium uptake and use[J]. Physiologia Plantarum, 2008, 133(4): 624–636. DOI: 10.1111/j.1399-3054.2008.01079.x
[5] Damon P M, Rengel Z. Wheat genotypes differ in potassium efficiency under glasshouse and field conditions[J]. Australian Journal of Agricultural Research, 2007, 58: 816–825. DOI: 10.1071/AR06402
[6] Zhang H, Rong H, Pilbeam D. Signalling mechanisms underlying the morphological responses of the root system to nitrogen in Arabidopsis thaliana[J]. Journal of Experimental Botany, 2007, 58(9): 2329–2338. DOI: 10.1093/jxb/erm114
[7] Trehan S P, Sharma R C. Potassium uptake efficiency of young plants of three potato cultivars as related to root and shoot parameters[J]. Communications in Soil Science &Plant Analysis, 2002, 33(11–12): 1813–1823.
[8] Wang T B, Gassmann W. Rapid up–regulation of HKT1, a high–affinity potassium transporter gene, in roots of barley and wheat following withdrawal of potassium[J]. Plant Physiology, 1998, 118: 651–659. DOI: 10.1104/pp.118.2.651
[9] Woodend J J, Glass A D M, Person C O. Genetic variation in the uptake and utilization of potassium in wheat (Triticumaestivum L.) varieties grown under potassium stress[J]. Genetic Aspects of Plant Mineral Nutrition, 1987, 27: 383–391.
[10] Flint–Garcia S A, Thornsberry J M, And E S, et al. Structure of linkage disequilibrium in plants[J]. Annual Review of Plant Biology, 2003, 54(4): 357–374.
[11] Flint–Garcia S A, Thuillet A C, Yu J, et al. Maize association population: a high-resolution platform for quantitative trait locus dissection [J]. The Plant Journal, 2005, 44(6): 1054–1064. DOI: 10.1111/j.1365-313X.2005.02591.x
[12] 武玉国, 吴承来, 秦保平, 等. 黄淮冬麦区175个小麦品种的遗传多样性及SSR标记与株高和产量相关性状的关联分析[J]. 作物学报, 2012, 38: 1–11. Wu Y G, Wu C L, Qin B P, et al. Diversity of 175 wheat varieties from yellow and huai river valleys facultative wheat zone and association of SSR markers with plant height and yield related traits[J]. Acta Agronomica Sinica, 2012, 38: 1–11.
[13] Zhang D L, Hao C Y, Wang L F, et al. Identifying loci influencing grain number by microsatellite screening in bread wheat (Triticumae stivum L.) [J]. Planta(Berlin), 2012, 236(5): 1507–1517.
[14] 张国华, 高明刚, 张桂芝, 等. 黄淮麦区小麦品种(系)产量性状与分子标记的关联分析[J]. 作物学报, 2013, 39: 1187–1199. DOI: 10.3724/SP.J.1006.2013.01187 Zhang G H, Gao M G, Zhang G Z, et al. Association analysis of yield traits with molecular markers in Huang-Huai river valley winter wheat region, China [J]. Acta Agronomica Sinica, 2013, 39: 1187–1199. DOI: 10.3724/SP.J.1006.2013.01187
[15] Kong F M, Guo Y, Liang X, et al. Potassium (K) effects and QTL mapping for K efficiency traits at seedling and adult stages in wheat[J]. Plant &Soil, 2013, 373(1–2): 877–892.
[16] 宫晓平. 小麦钾养分效率及产量相关性状的QTL分析[D]. 山东泰安: 山东农业大学硕士学位论文, 2014. Gong X P. QTL mapping for K efficiency traits at seedling and yield related traits at harvest stages in wheat[D]. Tai’an, Shandong: MS Thesis of Shandong Agricultural University, 2014.
[17] Siddiqi M Y, Glass A D M. Utilization index: A modified approach to the estimation and comparison of nutrient utilization efficiency in plants[J]. Journal of Plant Nutrition, 1981, 4(3): 289–302. DOI: 10.1080/01904168109362919
[18] Knapp S J, Stroup W W, Ross W M. Exact confidence intervals for heritability on a progeny mean basis[J]. Crop Science, 1985, 25(1): 192–194. DOI: 10.2135/cropsci1985.0011183X002500010046x
[19] Wang S, Wong D, Forrest K, et al. Characterization of polyploid wheat genomic diversity using a high-density 90, 000 single nucleotide polymorphism array[J]. Plant Biotechnology Journal, 2014, 12: 787–796. DOI: 10.1111/pbi.12183
[20] 屈春艳. 水旱条件下小麦产量性状和抗旱性的全基因组关联分析[D]. 山东泰安: 山东农业大学博士学位论文, 2018. Qu C Y. Genome-wide association study on yield traits and drought tolerance under irrigation and drought conditions in wheat[D]. Tai’an, Shandong: PhD Dissertation of Shandong Agricultural University, 2018.
[21] Pritchard J K, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data[J]. Genetics, 2000, 155(2): 945–959.
[22] Evanno G S, Regnaut S J, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study[J]. Molecular Ecology, 2005, 14(8): 2611–2620. DOI: 10.1111/j.1365-294X.2005.02553.x
[23] Hermans C, Hammond J, White P, et al. How do plants respond to nutrient shortage by biomass allocation?[J]. Trends in Plant Science, 2006, 11(12): 610–617. DOI: 10.1016/j.tplants.2006.10.007
[24] 钟晓兰, 张德远, 李江涛, 等. 施钾对烤烟钾素吸收利用效率及其产量和品质的影响[J]. 土壤, 2008, 40: 216–22. DOI: 10.3321/j.issn:0253-9829.2008.02.011 Zhong X L, Zhang D Y, Li J T, et al. Effects of K application on K absorption, K utilization efficiency, and yield and quality of flue-cured tobacco[J]. Soils, 2008, 40: 216–22. DOI: 10.3321/j.issn:0253-9829.2008.02.011
[25] Sukumaran S, Dreisigacker S, Lopes M, et al. Genome-wide association study for grain yield and related traits in an elite spring wheat population grown in temperate irrigated environments[J]. Theoretical and Applied Genetics, 2015, 128(2): 353–363. DOI: 10.1007/s00122-014-2435-3
[26] Shi W P, Hao C Y, Zhang Y, et al. A combined association mapping and linkage analysis of kernel number per spike in common wheat (Triticum aestivum L.)[J]. Frontiers in Plant Science, 2017, 8: 1412. DOI: 10.3389/fpls.2017.01412
[27] Zanke C D, Ling J, Plieske J, et al. Analysis of main effect QTL for thousand grain weight in European winter wheat (Triticum aestivum L.) by genome-wide association mapping[J]. Frontiers in Plant Science, 2015, 6: 644.
-
期刊类型引用(4)
1. 赵秋月,张咪咪,孙清鹏,韩俊. 小麦微核心种质株高及籽粒性状遗传分析. 安徽农业科学. 2022(07): 37-40+45 .  百度学术
百度学术
2. 刘硕,张慧,高志源,许吉利,田汇. 437个小麦品种钾收获指数的变异特征. 中国农业科学. 2022(07): 1284-1300 . 百度学术
3. 郭营,孙俊生,屈春艳,郭宝晋,赵岩,李斯深. 小麦黄花叶病抗性鉴定和全基因组关联分析. 山东农业科学. 2021(05): 51-57 . 百度学术
4. 郑福兴,颜安,高雪,严勇亮,王睿,耿洪伟. 水旱处理下小麦叶绿素相对含量全基因组关联分析. 植物遗传资源学报. 2021(05): 1334-1347 . 百度学术
其他类型引用(6)
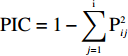
计量
- 文章访问数: 1892
- HTML全文浏览量: 1118
- PDF下载量: 53
- 被引次数: 10
